|
Biopolímeros (4831) |
 |
|
Módulo 4. Ficha
4.16
|
|
|
Biopolímeros (4831) |
|
|
Módulo 4. Ficha
4.16
|
ENZIMAS
4.8. Ejemplos de mecanismo catalítico enzimático: b-lactamasas y proteasas
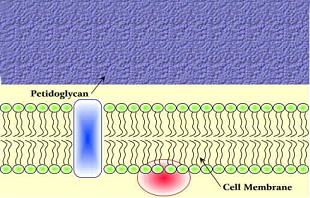
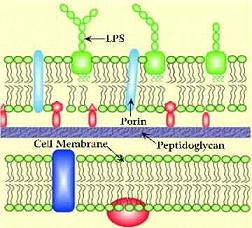
Las células bacterianas (procariotas) tienen una pared celular externa constituida por una red de peptidoglicanos que da forma a la bacteria y la protege del exterior. Dependiendo del tipo de organismo, la pared celular puede estar recubierta de una segunda membrana celular que regula la conexión entre el interior celular y el medio (Gram -) o no (Gram +).
En las bacterias gram +, la red de peptidoglicanos que constituyen la pared celular es totalmente externa. En las gram -, aún hay una membrana celular más externa, con canales selectivos (porinas) que pueden ser modificados para modular la selectividad a la entrada de nutrientes.
La red de péptidoglicanos está compuesta por largas cadenas de polisacáridos, formadas con unidades de ácido N-acetilmurámico y N-acetilglucosamina, que se entrecruzan mediante la unión a pequeñas unidades peptídicas. Las bacterias tienen toda una batería de enzimas que ayudan a la formación de la pared bacteriana, entre ellos hay DD-peptidasas (carboxipeptidasas y traspeptidasas) que son los targets de los antibióticos b-lactámicos.
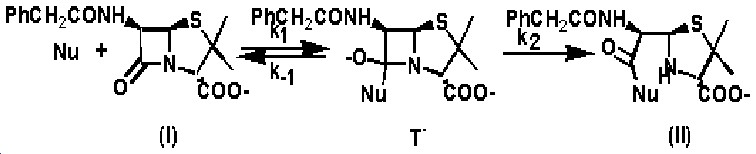
La disposición espacial de los átomos y la distribución electrónica en el residuo D-alanil-D-alanina, izquierda, es idéntica a la de los átomos del anillo b-lactámicos, derecha. Estos enzimas bacterianos son serin-enzimas y confunden a su substrato natural, residuos D-alanil-D-alanina, con el anillo b-lactámico, de manera que se unen a él y forman el complejo de Michaelis (EC) que evoluciona hacia la formación de un acil-enzima (EC*) de manera similar a como lo hacen con el sustrato. En el intermedio acil-enzima, el antibiótico rompe su enlace lactámico de forma que el sistema no puede evolucionar y el enzima queda irreversiblemente inhibido.
Muchas bacterias han desarrollado estrategias para luchar contra la acción de los antibióticos betalactámicos; la producción de b-lactamasas es una de ellas. Las b-lactamasas son enzimas que catalizan la hidrólisis del anillo b-lactámico. Muchas de ellas son también serin-enzimas y provienen de las DD-peptidasas por evolución. Respecto a estas, han modificado su estructura de forma que, por un lado han acomodado una molécula de agua en el sitio activo, la cual es utilizada posteriormente para la hidrólisis del acil-enzima y por otro, han modificado el sitio activo para que no pueda unir el peptido del entrecruzamiento y evita así su hidrólisis.
Las b-lactamasas aceleran la formación del intermedio tetraédrico acomodando el oxianión generado en un entorno con dos fuertes dipolos, N-H, que interaccionan electrostáticamente con él.
|
|
||||
|
dominio a/b |
dominio a | nucleófilo | "oxianion hole" | centro activo |
|
||||
|
|
||||
|
Nucleófilo |
O1 carboxilato
|
"Oxianion hole"
|
O2 carboxilato
|
|
|
||||
La rotura del enlace CN de la betalactama, implica la cesión de un protón al N saliente. En el proceso, deben estar implicados la Tyr150 y la Lys67. En la segunda etapa del proceso catalítico, una molécula de agua debe ser activada como nucleófilo, posiblemente por la Lys67, para atacar al carbono carbonílico del acil-enzima y comenzar el proceso de desacilación. El sitio electrófilo "oxianión hole" sigue siendo el mismo.
El mecanismo descrito anteriormente es prácticamente universal para todos los enzimas que catalizan la rotura hidrolítica de enlaces amida o esteres. En la figura inferior se muestra el mecanismo catalítico de la hidrólisis del enlace peptídico en proteínas catalizado por una proteasa, la quimotripsina.
Mecanismo de hidrólisis del enlace peptídico catalizado por quimiotripsina. En el complejo enzima-substrato (ES) el oxígeno carbonílico está anclado por el extremo positivo de los dipolos HN de la cadena polipeptídica. El carbono carbonílico está así anclado en la posición correcta para que se produzca el ataque nucleófilo del hidroxilo de la Ser-195 y la His-57 actúa como catalízador básico general activando el g-OH del nucleófilo. En el primer intermedio tetrahédrico (IT1) Se desarrolla una carga negativa sobre el oxígeno que es estabilizada por los dipolos de la cadena. La interacción entre el Asp-102 y la His-57 aumenta para estabilizar la carga positiva que se desarrolla en ella. Esta, a su vez activa el grupo R'NH- como grupo saliente mediante la cesión de un protón. El mecanismo de desacilación ocurre mediante la activación como nucleófilo de la molécula de agua por la His-57, la generación del segundo intermedio tetrahédrico (IT2) cuya rotura regenera la Ser-195 libre y los productos quedan aún anclados en el complejo enzima-productos.(EP).
A
Docencia  |